
2020
W poznaniu struktur i procesów komórkowych organizmów pro i eukariotycznych ogromną rolę odegrały organizmy modelowe. Celem ich zastosowania było nie tyle uzyskanie wiedzy o konkretnym procesie przebiegającym w danym organizmie, ale zrozumienie ogólnych zasad funkcjonowania organizmów żywych, w tym również organizmu człowieka.
 Do organizmów modelowych zalicza się: bakterie (E. coli), rzodkiewnik pospolity (Arabidpopsis thalina), nicień (Caenorhabditis elegans), muszkę owocową (Drosophila melanogaster), zeberkę (Taenipygia guttata), mysz (Mus musculus) oraz drożdże (w tym Saccharomyces cerevisiae).
Do organizmów modelowych zalicza się: bakterie (E. coli), rzodkiewnik pospolity (Arabidpopsis thalina), nicień (Caenorhabditis elegans), muszkę owocową (Drosophila melanogaster), zeberkę (Taenipygia guttata), mysz (Mus musculus) oraz drożdże (w tym Saccharomyces cerevisiae).
Drożdże tego gatunku, to grzyby z klasy Ascomycetes (workowców), a więc organizmy o budowie eukariotycznej. Występują one pospolicie w wielu środowiskach naturalnych takich jak: gleba, powierzchnia roślin (w zbiorowiskach leśnych szczególnie upodobały sobie korę dębów) oraz bogate w cukry proste – kwiaty i owoce. Wykorzystywane nieświadomie przez człowieka od tysiącleci do spulchniania pieczywa oraz w produkcji win i piwa są one traktowane jako pierwszy „udomowiony” organizm. Obecnie, wyselekcjonowane i „ulepszone” szczepy S. cerevisiae są wykorzystywane na skalę przemysłową m.in. w produkcji preparatów farmaceutycznych oraz w przemyśle winiarskim, piwowarskim, gorzelniczym oraz przy wypieku pieczywa.
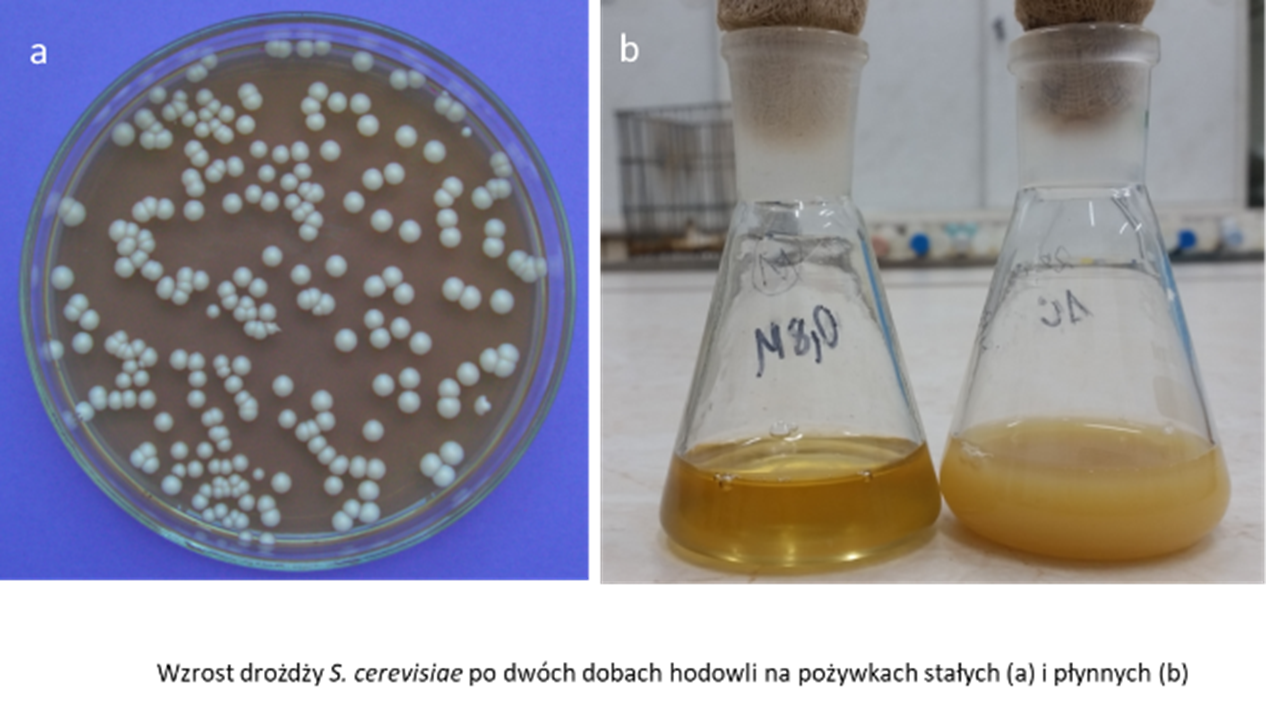
Zajmują one także ważną pozycję w badaniach naukowych. Jako organizmy eukariotyczne charakteryzują się podobieństwem organizacji struktur komórkowych i procesów biochemicznych do komórek wyższych eukariota, w tym ssaków. Zjawisko to wynika ze znacznego konserwatyzmu ewolucyjnego struktur i funkcji komórkowych. Jako jednokomórkowe mikroorganizmy można je łatwo hodować w warunkach laboratoryjnych na standardowych podłożach płynnych i stałych. Namnażają się szybko (ich cykl podziałowy w optymalnych warunkach wynosi około 90 minut). Dzięki temu już po dwóch dobach od zaszczepienia pożywek można analizować wzrost tych mikroorganizmów (rys.1). Mogą one występować w formie stabilnych linii haploidalnych lub diploidalnych, które mogą rozmnażać się wegetatywnie poprzez proces specyficzny dla tego gatunku- pączkowanie. Są względnymi tlenowcami. Hodowane w warunkach tlenowych przy stosunkowo niskim stężeniu glukozy zdobywają energię na drodze oddychania tlenowego, natomiast przy dużym stężeniu glukozy (2% i więcej) lub w warunkach beztlenowych – za pomocą fermentacji. Pomimo tego, że jest to proces mniej wydajny energetycznie to jest on preferowanym sposobem zdobywania energii u tego mikroorganizmu (efekt Crabtree). Pod tym względem drożdże te przypominają niektóre typy komórek ssaczych takie jak np. komórki nowotworowe 1. Ważną cechą drożdży wynikającą z ich specyficznego metabolizmu energetycznego (zdolność do oddychania tlenowego lub fermentacji) jest możliwość funkcjonowania bez aktywnych mitochondriów. Komórki z uszkodzonymi, nieaktywnymi mitochondriami tzw. „mutanty petite” pojawiają się spontanicznie (z małą częstotliwością) lub też można je uzyskać na drodze mutacji. Są one ciekawym obiektem badań, gdyż tego typu mutacje u organizmów wyższych są letalne.
Zaletą drożdży jest także to, że są one najlepiej scharakteryzowanym pod względem genetycznym organizmem. Genom jądrowy drożdży był pierwszym całkowicie zsekwencjonowanym genomem. Ponadto komórki drożdży łatwo poddają się manipulacjom genetycznym, co przyczyniło się do uzyskania licznych mutantów. Nie bez znaczenia jest także fakt, że są one bezpieczne dla badaczy (nie wytwarzają toksycznych metabolitów) i są nieszkodliwe dla środowiska. Badania na drożdżach z gatunku S. cerevisiae umożliwiły poznanie uniwersalnych dla organizmów eukariotycznych procesów takich jak np.: cykl komórkowy i jego regulacja, transkrypcja, transport pęcherzykowy i autofagia, biologia telomerów, sygnalizacja uszkodzeń DNA i mechanizmy jego naprawy, proces starzenia, metabolizm glukozy, odpowiedź na stres środowiskowy czy biogeneza struktur subkomórkowych takich jak: mitochondriów, retikulum endoplazmatycznego, aparat Golgi’ego, wakuole i peroksysomy 2, 3, 4, 5, 6.
ul. Akademicka 13, 20-950 Lublin
NIP 712 010 37 75
REGON 000001896
e-Doręczenia: AE:PL-92700-40162-VCRBJ-25
ePUAP: /UP-Lublin/SkrytkaESP